




O sistema endócrino coordena as funções entre os diferentes órgãos por meio dos hormônios, que são substâncias químicas liberadas na corrente sanguínea por células específicas dentro de glândulas endócrinas (sem ductos). Uma vez na circulação, os hormônios afetam a função de tecidos alvo, que podem ser outra glândula endócrina ou órgão final. Alguns hormônios atuam nas células do órgão do qual foram liberados (efeito parácrino), alguns inclusive no mesmo tipo de célula (efeito autócrino).
Hormônios podem ser
Peptídeos (um ou mais aminoácidos conectados por ligações químicas) de vários tamanhos
Esteroides (derivados do colesterol)
Derivados de aminoácidos
Os hormônios se ligam seletivamente a receptores localizados na superfície ou dentro das células-alvo. Os receptores no interior das células interagem com hormônios que regulam a função gênica (p. ex., corticoides, vitamina D, hormônio tireoidiano). Os receptores na superfície das células se ligam a hormônios que regulam atividades enzimáticas ou afetam canais de íons (p. ex., GH, TRH).
Doenças endócrinas resultam de rupturas das glândulas endócrinas e/ou seus tecidos alvo.
Hipófise e seus órgãos-alvo

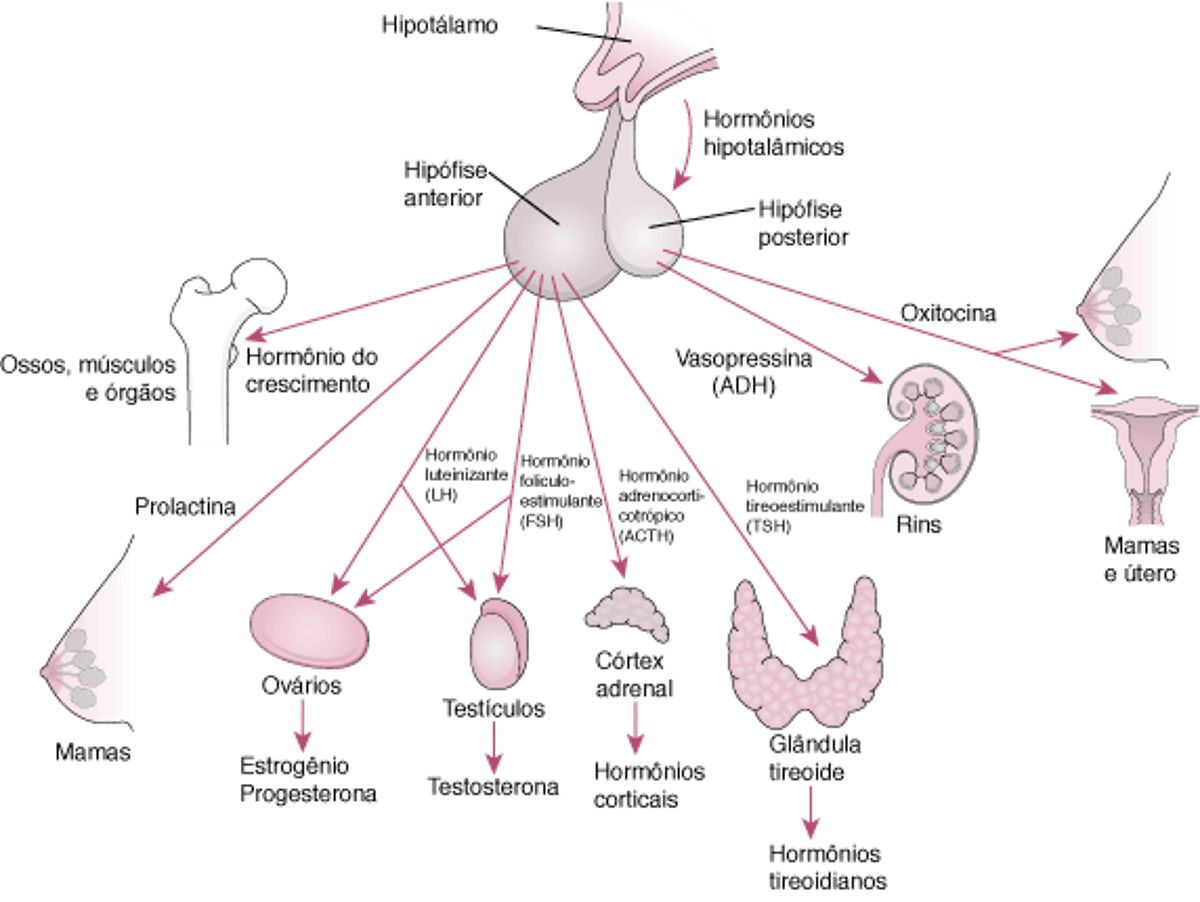
Relações hipotalâmico-hipofisárias
As funções dos órgãos endócrinos periféricos são controladas, em vários graus, pelos hormônios hipofisários. Algumas funções (p. ex., secreção de insulina pelo pâncreas, primariamente controlado pelo nível de glicose no sangue) são controlados em uma extensão mínima ou são independentes do controle da hipófise (p. ex., secreção de hormônio paratireoideo pelas glândulas paratireoides, principalmente em resposta aos níveis de cálcio no sangue), ao passo que muitos (p. ex., secreção de hormônios tireoidianos ou gonadais) são controlados em grande medida. A secreção de hormônios hipofisários é controlada pelo hipotálamo.
A interação entre o hipotálamo e a hipófise (chamado eixo hipotálamo-hipófise) é um sistema de controle de feedback. O hipotálamo recebe mensagens virtuais de todas as outras áreas do sistema nervoso central e as utiliza para enviar mensagens para a hipófise. Em resposta, a hipófise libera vários hormônios que estimulam certas glândulas endócrinas do corpo. As alterações nas concentrações circulantes de hormônios produzidas pelas glândulas endócrinas são detectadas pelo hipotálamo, que então aumenta ou diminui seu estímulo sobre a hipófise para manter a homeostase.
O hipotálamo modula as atividades dos lobos anterior e posterior da hipófise de maneiras diferentes. Os neuro-hormônios sintetizados no hipotálamo alcançam a hipófise anterior (adeno-hipófise) por meio de um sistema portal vascular especializado e regulam a síntese e a liberação de 6 hormônios peptídeos principais da adeno-hipófise (ver figura Hipófise e seus órgãos alvo). Esses hormônios da adeno-hipófise regulam as glândulas endócrinas periféricas (tireoide, adrenais e gônadas), assim como crescimento e lactação. Não há conexões neurais diretas entre o hipotálamo e a adeno-hipófise.
Por outro lado, a hipófise posterior (neuro-hipófise) contém axônios originários de corpos de neurônios localizados no hipotálamo. Esses axônios servem como locais de armazenagem para 2 hormônios peptídicos, vasopressina (hormônio antidiurético) e ocitocina, sintetizados no hipotálamo; esses hormônios atuam na periferia para regular o equilíbrio hídrico, a ejeção de leite e as contrações uterinas.
Virtualmente todos os hormônios produzidos pelo hipotálamo e pela hipófise são liberados de forma pulsátil; períodos de liberação são intercalados com períodos de inatividade. Alguns hormônios (p. ex., ACTH, GH, prolactina) apresentam ritmos circadianos definidos; outros (p. ex., LH e FSH durante o ciclo menstrual) apresentam ritmos mensais com ritmos circadianos sobrepostos.
Controles hipotalâmicos
Até agora, foram identificados 7 neuro-hormônios hipotalâmicos (ver tabela Neuro-hormônios hipotalâmicos). Exceto a amina biogênica dopamina, todos são peptídeos pequenos. Vários são produzidos perifericamente, assim como no hipotálamo, e atuam em sistemas parácrinos locais, em especial no trato gastrointestinal. O peptídeo intestinal vasoativo, que também estimula a liberação de prolactina, é um deles.
Os neuro-hormônios podem controlar a liberação de múltiplos hormônios hipofisários. A regulação da maioria dos hormônios da adeno-hipófise depende de sinais estimulantes do hipotálamo; apenas a prolactina é regulada por estímulos inibitórios. Se a haste hipofisária (que conecta a hipófise ao hipotálamo) for lesionada, a liberação de prolactina aumenta, ao passo que a liberação dos outros hormônios da adeno-hipófise diminui.
Várias anormalidades hipotalâmicas (incluindo tumores, encefalite e outras lesões inflamatórias) podem alterar a liberação dos neuro-hormônios hipotalâmicos. Como os neuro-hormônios são sintetizados em diferentes centros no hipotálamo, algumas doenças afetam apenas um neuropeptídeo e outras afetam vários. O resultado pode ser diminuição ou aumento da secreção de neuro-hormônios. As síndromes clínicas que resultam de disfunções de hormônios hipofisários (p. ex., diabetes insípido, acromegalia e hipo-hipofisarismo) são discutidas em outras partes.
Função da adeno-hipófise
As células do lobo anterior (que constituem 80% do peso da hipófise) sintetizam e liberam hormônios necessários para o crescimento e desenvolvimento normais e estimulam a atividade de várias glândulas-alvo.
Hormônio adrenocorticotrófico (ACTH)
Também conhecido como corticotrofina. O hormônio liberador de corticotrofina (CRH) é o principal estimulador da liberação de ACTH, mas a vasopressina desempenha um papel durante o estresse. O ACTH induz a liberação, pelo córtex adrenal, de cortisol e vários andrógenos fracos, como sulfato de desidroepiandrosterona (DHEAS). O cortisol e outros corticoides circulantes (inclusive os corticoides exógenos) inibem a liberação de CRH e ACTH. O eixo CRH-ACTH-cortisol é um componente central da resposta ao estresse. Sem o ACTH, ocorre atrofia do córtex adrenal e a liberação de cortisol virtualmente cessa.
Hormônio estimulante da tireoide (TSH)
Regula a estrutura e a função da tireoide e estimula a síntese e liberação de hormônios tireoidianos. A síntese e liberação de TSH são estimuladas pelo hormônio hipotalâmico TRH e suspensas (por feedback negativo) pelos hormônios tireoidianos circulantes.
Hormônio luteinizante (LH) e hormônio estimulante de folículos (FSH)
Controlam a produção de hormônios sexuais. A síntese e liberação de LH e FSH são estimuladas principalmente pelo GnRH e suprimidas por estrogênio e testosterona. Um fator controlador da liberação de GnRH é kisspeptina, um peptídeo hipotalâmico que é desencadeado pelo aumento dos níveis de leptina na puberdade. Dois hormônios gonadais, ativina e inibina, afetam apenas o FSH; a ativina é estimulante, e a inibina é inibidora.
Em mulheres, LH e FSH estimulam o desenvolvimento dos folículos ovarianos e a ovulação.
Em homens, o FSH atua nas células de Sertoli e é essencial para a espermatogênese; o LH atua sobre as células de Leydig dos testículos para estimular a biossíntese de testosterona.
Hormônio do crescimento (GH)
Estimula o crescimento somático e regula o metabolismo. O GHRH é o principal estimulante e a somatostatina é o principal inibidor da síntese e liberação do GH. O GH controla a síntese do fator de crescimento semelhante à insulina 1 (IGF-1, também chamado de somatomedina-C), que em grande medida controla o crescimento. Embora o IGF-1 seja produzido em muitos tecidos, o fígado é a principal fonte. Existe uma variante de IGF-1 no músculo, onde atua aumentando a força muscular. Essa variante está menos sob controle do GH do que a variante hepática.
Os efeitos metabólicos do GH são bifásicos. O GH exerce, inicialmente, efeitos semelhantes à insulina, aumentando a captação de glicose nos tecidos muscular e adiposo, estimulando a captação de aminoácidos e a síntese proteica no fígado e nos músculos e inibindo a lipólise no tecido adiposo. Várias horas mais tarde, ocorrem efeitos metabólicos mais profundos do tipo anti-insulina. Eles incluem a inibição da captação e utilização de glicose, causando elevação da glicemia e lipólise no sangue, o que aumenta a quantidade de ácidos graxos livres no plasma. As concentrações de GH aumentam no jejum, mantendo as concentrações de glicose e mobilizando gordura como forma alternativa de combustível metabólico. A produção de GH diminui com o envelhecimento. A grelina, um hormônio produzido pela região do fundo do estômago, promove a liberação de GH da hipófise, aumenta a ingestão de alimentos e em modelos animais melhora a memória.
Prolactina
É produzida por células denominadas lactotrofos, que constituem cerca de 30% das células da adeno-hipófise. A hipófise dobra de tamanho durante a gestação, em grande parte em decorrência da hiperplasia e hipertrofia dos lactotrofos. Em seres humanos, a principal função da prolactina é estimular a produção de leite. A liberação de prolactina também ocorre durante a atividade sexual e o estresse. A prolactina pode ser um indicador sensível de disfunção hipofisária; é o hormônio que, com mais frequência, é produzido em excesso pelos tumores hipofisários, e geralmente é o primeiro hormônio que se torna deficiente em razão de doenças infiltrativas ou compressão por tumor da hipófise.
Outros hormônios
Vários outros hormônios são produzidos pela glândula hipófise anterior. Estes incluem pró-opiomelanocortina (POMC, que dá origem ao ACTH), alfa e beta-hormônio estimulante do melanócito (MSH), beta-lipotrofina (β-LPH), as encefalinas e as endorfinas. POMC e MSH podem causar hiperpigmentação da pele e têm significado clínico apenas nas doenças com elevação de ACTH (p. ex., doença de Addison, síndrome de Nelson). A função da β-LPH é desconhecida. Encefalinas e endorfinas são opioides exógenos que se ligam e ativam receptores opioides ao longo do sistema nervoso central.
Função da neuro-hipófise
A hipófise posterior libera a vasopressina (também chamada vasopressinaarginina ou hormônio antidiurético ADH) e a ocitocina. Ambos são liberados em resposta a impulsos neurais e têm meia-vida de aproximadamente 10 minutos.
Vasopressina (hormônio antidiurético, ADH)
A vasopressina age primariamente para promover a conservação de água pelos rins aumentando a permeabilidade do epitélio tubular distal à água. Em altas concentrações, a vasopressina causa vasoconstrição. Assim como a aldosterona, a vasopressina desempenha um papel importante na manutenção da homeostase líquida e da hidratação vascular e celular. O principal estímulo para liberação da vasopressina é o aumento da pressão osmótica da água no corpo, detectado pelos osmorreceptores no hipotálamo.
Outro estímulo importante é a depleção de volume, detectada por barorreceptores no átrio esquerdo, veias pulmonares, seio carotídeo e arco aórtico e depois transmitidos para o sistema nervoso central pelos nervos vago e glossofaríngeo. Outros estímulos para a liberação da vasopressina incluem dor, estresse, êmese, hipóxia, atividade física, hipoglicemia, agonistas colinérgicos, betabloqueadores, angiotensina e prostaglandinas. Os inibidores da liberação de vasopressina incluem álcool, alfa-bloqueadores e glicocorticoides.
A falta de vasopressina causa diabetes insípido central. A incapacidade dos rins para responder normalmente à vasopressina causa diabetes insípido nefrogênico. A remoção da glândula hipófise habitualmente não resulta em diabetes insípido permanente porque alguns neurônios hipotalâmicos remanescentes produzem pequenas quantidades da vasopressina.
Copeptina é coproduzida com vasopressina na neuro-hipófise. Sua medida pode ser útil para diferenciar as causas de hiponatremia.
Ocitocina
A ocitocina tem 2 alvos importantes:
As células mioepiteliais das mamas, que circundam os alvéolos da glândula mamária
Células do músculo liso do útero
A sucção estimula a produção de ocitocina, que causa contração das células mioepiteliais. Essa contração causa a movimentação do leite dos alvéolos para os seios maiores para ser ejetado (isto é, o reflexo de saída de leite das mães que estão amamentando). A ocitocina estimula as contrações uterinas das células musculares lisas e a sensibilidade uterina à ocitocina aumenta durante a gestação. Entretanto, as concentrações plasmáticas não aumentam abruptamente durante o parto e o papel da ocitocina na deflagração do trabalho de parto não foi elucidado.
Não há estímulo reconhecido para a liberação de ocitocina em homens, embora estes apresentem concentrações extremamente baixas.







