ニューロンは活動電位を発生させて,これを軸索に沿って伝導した後,このシグナルを神経伝達物質の放出によりシナプスを介して伝達し,別のニューロンまたは効果細胞(例,筋細胞,外分泌および内分泌細胞の大半)の応答を誘発する。放出された神経伝達物質は別のニューロンの受容体に結合する。神経伝達物質を放出するニューロンは,シナプス前ニューロンと呼ばれる。神経伝達物質によるシグナルを受け取るニューロンは,シナプス後ニューロンと呼ばれる。関与する神経伝達物質と受容体の関係に応じて,受容細胞はシグナルによって刺激される場合と抑制される場合がある。
ときに,ニューロン間のシグナル伝達が逆方向に起こる場合がある(逆行性神経伝達と呼ばれる)。その場合,シナプス後ニューロンの樹状突起(シグナルを受け取るニューロンの枝)から神経伝達物質が放出され,それがシナプス前ニューロンの受容体に作用する。逆行性伝達により,シナプス前ニューロンからのさらなる神経伝達物質の放出を阻害することができ,ニューロン間の活動やコミュニケーションの水準を制御するのに役立っている。
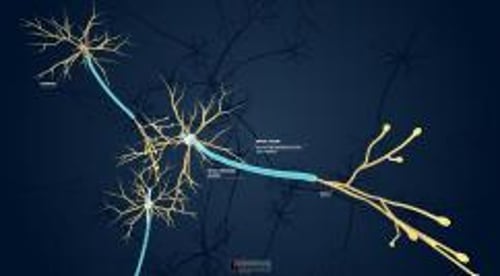
中枢神経系では,相互接続が複雑である。あるニューロンから別のニューロンへのインパルスは以下の経路をたどる可能性がある:
軸索から細胞体へ
軸索から樹状突起へ
細胞体から細胞体へ
樹状突起から樹状突起へ
ニューロンは,複数のニューロンから同時に多くの興奮性および抑制性インパルスを受け取り,同時に発生したインパルスを様々な発火パターンに統合することができる。
伝導
軸索に沿った活動電位の伝導は電気的な現象であり,軸索膜を介したナトリウムとカリウムの交換によって引き起こされる。個々のニューロンは,毎回の刺激後に同じ大きさの活動電位を発生させ,それを軸索に沿って一定の速度で伝導する。伝導速度は軸索の直径と髄鞘化の程度に依存し,細い無髄線維では1~4m/秒,太い有髄線維では75m/秒である。伝導速度は有髄線維の方が速いが,これは有髄線維には一定の間隔で軸索が露出した部分(ランビエ絞輪)があるためである。電気パルスは,髄鞘化された軸索部分を飛び越えて,絞輪から絞輪へと跳躍して伝導していく。したがって,髄鞘に影響を及ぼす疾患(例,多発性硬化症)ではインパルスの伝導が障害され,それにより様々な神経症状が引き起こされる。
伝達
インパルスの伝達は化学的な現象であり,神経終末(末端)から特異的な神経伝達物質が放出することにより引き起こされる。神経伝達物質はシナプス間隙を越えて拡散し,隣接するニューロンまたは効果細胞の特異的受容体に短時間結合する。結合する受容体に応じて,生じる反応は興奮性または抑制性となる。通常,ニューロンは互いに接触することなく,代わりにシナプスを介した神経伝達物質の伝達によってコミュニケーションが行われる。
ある種のシナプス(電気シナプス)では,神経伝達物質は関与せず,イオンチャネルによってシナプス前後のニューロンの細胞質が直接連結される。この形態の伝達が最も速く進行する。
神経細胞体は,ほとんどの神経伝達物質を合成する酵素を産生しており,神経伝達物質は神経終末の小胞内に貯蔵される(神経伝達の図を参照)。1つの小胞内にある量(通常は分子数で数千)を1量子(quantum)と呼ぶ。神経終末に到達した膜活動電位によって軸索のカルシウムチャネルが開口すると,カルシウムイオンが流入する結果,小胞膜が神経終末膜に融合することにより,多数の小胞から神経伝達物質の分子が放出される。膜融合により出口ができると,そこから開口分泌によってシナプス間隙に分子が放出される。
興奮性および抑制性シグナル
神経伝達物質の放出によって引き起こされる反応は,シナプス後ニューロンを刺激または活性化することもあれば,その活動を阻害または遮断することもある。シナプス後ニューロンは,多くのニューロンから複数の神経伝達物質シグナルと電気信号を受け取る。それらを受け取ったニューロンは最終的にこれらの入力情報を合算するが,より多くの興奮性シグナルを受け取ると,そのニューロンが発火して他のニューロンにシグナルを伝達する。シグナルの合算値が抑制的であれば,そのニューロンは発火せず,他のニューロンの活動に影響を及ぼさない。このように反応を合算することを加重と呼ぶ。
加重にはほかに以下の2種類がある:
空間的加重:1つのニューロンが異なる場所で複数のインパルスを受け取ると,ニューロンがそれらを合算する
時間的加重:短時間のうちにインパルスが受け取られると,それらが合算される
ニューロンがシグナルを生成して発火するには,閾値電位に到達する必要がある。閾値電位は,ナトリウムおよびカリウムイオンの交換中に細胞内に流入するナトリウムの正味の増加量によって生成される。十分なナトリウムが細胞内に流入すると,閾値に到達するが,閾値に到達すると,活動電位が発火し,それがニューロンの膜に沿って移動する。閾値に到達しなければ,活動電位は発生しない。
神経伝達
活動電位により軸索のCaチャネルが開口する(図には示していない)。Ca2+が神経伝達物質(NT)の貯蔵場所である小胞からのNTの放出を活性化する。NTの分子でシナプス間隙が満たされる。一部がシナプス後受容体と結合して,応答を惹起する。それ以外のNTは,軸索内に回収されるか,周囲の組織中に貯蔵されたり拡散したりする。 |

神経終末における神経伝達物質の量は,基本的には神経の活動に依存せず,神経伝達物質前駆体の取込みの調節や神経伝達物質の合成または分解に関与する酵素の活性の調節によって,比較的一定に保たれている。シナプス前受容体を刺激するとシナプス前部での神経伝達物質の合成を減少させ,シナプス前受容体を遮断すると合成を増加させることができる。
神経伝達物質の持続的な活動を遮断するおよび/または受容体の急速かつ反復的な活性化を可能にするには,神経伝達物質と受容体との相互作用は速やかに終結する必要がある。受容体と相互作用を起こした神経伝達物質には,以下のいずれかが起こる:
能動的かつATP依存的にシナプス前神経終末に回収され(再取り込み),リサイクルまたは破壊される。
受容体付近の酵素によって除去される。
周辺の領域に拡散して除去される。
神経終末に取り込まれた神経伝達物質は,軸索終末の顆粒内または小胞内に再び詰め込まれ,再利用される。
これらのプロセスに機能障害が起こると,臨床的な疾患の発生につながる可能性がある。例えば,アルツハイマー病における記憶障害には,新たな記憶の構築を媒介するシナプス内の神経伝達物質であるアセチルコリンの不足が関連していると考えられている。特定の薬剤(例,ドネペジル,ガランタミン,リバスチグミン)は,アセチルコリンエステラーゼという(アセチルコリンを分解する)酵素を阻害し,シナプスにおけるアセチルコリンの量を増加させる。その結果,記憶機能が改善することがある。
ある種のニューロンは,単独で2つ以上の神経伝達物質(例えば,アセチルコリンとグルタミン酸)を放出できる(共伝達[cotransmission]と呼ばれる)。複数の神経伝達物質は,単一のシナプス後ニューロンに作用することもあれば,複数のシナプス後ニューロンに作用することもある。共伝達により,ニューロン間の複雑なコミュニケーションが可能になり,中枢神経系および末梢神経系(PNS)の様々なイベントを制御できる。
受容体
神経伝達物質の受容体は,細胞膜に広く分布するタンパク質複合体である。その性質によって,結合した神経伝達物質が興奮性に働くか,抑制性に働くかが決定される。神経伝達物質または薬物によって絶えず刺激されている受容体は脱感作(ダウンレギュレーション)を起こし,神経伝達物質による刺激を受けないか,薬物により慢性的に遮断されている受容体は感受性亢進(アップレギュレーション)を起こす。受容体のダウンレギュレーションやアップレギュレーションは,耐性や身体依存の発生に強く影響する。これらの概念は臓器または組織移植の際に特に重要であり,これらの状況では,脱神経のために受容体が神経伝達物質から遮断されている。離脱症状は,受容体の結合性または密度が変化したことによる反跳現象によって,少なくとも一部は説明できる。
大部分の神経伝達物質は主としてシナプス後受容体と相互作用するが,シナプス前ニューロンに発現する受容体もあり,神経伝達物質の放出を微調整している。
イオンチャネル型受容体と呼ばれる受容体ファミリー(例,N-メチル-d-グルタミン酸受容体,カイニン酸-キスカル酸受容体,ニコチン性アセチルコリン受容体,グリシン受容体,γ-アミノ酪酸[GABA]受容体)は,神経伝達物質と結合することで開口するイオンチャネルで構成されており,非常に迅速な応答に関与する。代謝型受容体と呼ばれる別の受容体ファミリー(例,セロトニン受容体,αおよびβアドレナリン受容体,およびドパミン受容体)では,神経伝達物質はGタンパク質と相互作用して,タンパク質のリン酸化やカルシウムの動員を介した連鎖反応を触媒する別の分子(cAMPなどのセカンドメッセンジャー)を活性化する;セカンドメッセンジャーが介在する細胞の変化は比較的緩徐であり,神経伝達物質に対するイオンチャネルによる迅速な応答を微細に調整することができる。セカンドメッセンジャーを活性する神経伝達物質より,特異的な受容体を活性する神経伝達物質の方がはるかに多い。
主要な神経伝達物質および受容体
少なくとも100種類の物質が神経伝達物質として作用し,そのうち約18種類が特に重要である。いくつかは若干異なる形態で存在する。神経伝達物質は以下のような複数のクラスに分類できる:
低分子(例,グルタミン酸,γ-アミノ酪酸,グリシン,アデノシン,アセチルコリン,セロトニン,ヒスタミン,ノルアドレナリン)
神経ペプチド(例,エンドルフィン)
ガス分子(例,一酸化窒素,一酸化炭素)
エンドカンナビノイド
グルタミン酸およびアスパラギン酸
これらのアミノ酸は,中枢神経系における主な興奮性神経伝達物質である。大脳皮質,小脳,および脊髄に存在する。ニューロン内では,グルタミン酸に反応して一酸化窒素(NO)合成が増加する。過剰なグルタミン酸は毒性を示すことがあり,細胞内カルシウム濃度,フリーラジカル,およびプロテアーゼ活性を増加させる。これらの神経伝達物質は,オピオイド治療に対する耐性の一因となったり,痛覚過敏に関与したりすることがある。
グルタミン酸受容体はNMDA(N-メチル-d-アスパラギン酸)受容体と非NMDA受容体に分類される。フェンシクリジン(PCP,エンジェルダストとしても知られる)とメマンチン(アルツハイマー病の治療に使用される)はNMDA受容体に結合する。
γ-アミノ酪酸
γ-アミノ酪酸(GABA)は脳における主な抑制性神経伝達物質である。グルタミン酸に由来するアミノ酸であり,グルタミン酸がグルタミン酸脱炭酸酵素により脱炭酸化されて生じる。受容体との相互作用の後,GABAは神経終末に能動的に回収されて代謝される。グリシンはその作用面でGABAに類似するが,主として脊髄の介在ニューロン(Renshaw細胞)や拮抗筋を弛緩させる回路に存在する。
GABA受容体はGABA-A(塩素イオンチャネルを活性化する)とGABA-B(cAMP合成を増強する)に分類される。GABA-A受容体は,ベンゾジアゼピン系薬剤,バルビツール酸系薬剤,ピクロトキシン,ムシモールなど,いくつかの神経刺激薬の作用部位である。アルコールもGABA-A受容体に結合する。GABA-B受容体は,筋攣縮(例,多発性硬化症において)の治療に使用されるバクロフェンにより活性化される。
セロトニン
セロトニン(5-ヒドロキシトリプタミン,5-HT)は,縫線核と橋および上位脳幹の正中に存在するニューロンにより産生される。トリプトファンはトリプトファン水酸化酵素によって5-ヒドロキシトリプトファンへと水酸化され,さらに脱炭酸化されてセロトニンとなる。セロトニン濃度は,トリプトファンの取込みとニューロン内のセロトニンを分解するモノアミン酸化酵素(MAO)によって調節される。最終的に,セロトニンは5-ヒドロキシインドール酢酸(5-HIAA)として尿中に排出される。
セロトニン(5-HT)受容体(少なくとも15のサブタイプがある)は,5-HT1(4つのサブタイプがある),5-HT2,および5-HT3に分類される。選択的セロトニン受容体作動薬(例,スマトリプタン)は片頭痛を抑える効果がある。
アセチルコリン
アセチルコリンは,延髄脊髄の運動ニューロン,自律神経節前線維,コリン作動性節後(副交感神経)線維,および中枢神経系の多くのニューロン(例,基底核,運動皮質)の主要な神経伝達物質である。アセチルコリンは,コリンとアセチルコエンザイムAからコリンアセチルトランスフェラーゼにより合成され,その作用は,局所におけるアセチルコリンエステラーゼによるコリンと酢酸への加水分解によって急速に終結する。アセチルコリン濃度は,コリンアセチルトランスフェラーゼとコリンの取込みによって調節されている。アルツハイマー病患者では,この神経伝達物質の濃度が低下している。
アセチルコリン受容体は,ニコチン性のN1(副腎髄質と自律神経節)およびN2(骨格筋)と,ムスカリン性のM1~M5(中枢神経系に広く分布)に分類される。M1受容体は自律神経系,線条体,大脳皮質,および海馬に,M2受容体は自律神経系,心臓,腸管平滑筋,後脳,および小脳に存在する。
ドパミン
ドパミンは,一部の末梢神経線維と多くの中枢ニューロン(例,黒質,中脳,腹側被蓋野,視床下部)の受容体と相互作用を起こす。アミノ酸のチロシンがドパミン作動性ニューロンに取り込まれ,チロシン水酸化酵素によって3,4-ジヒドロキシフェニルアラニン(ドパ)に変換され,これが芳香族l-アミノ酸脱炭酸酵素によって脱炭酸化されてドパミンとなる。放出されて受容体と相互作用した後,ドパミンは神経終末に能動的に回収される(再取り込み)。チロシン水酸化酵素とMAO(ドパミンを分解する)が神経終末におけるドパミン濃度を調節している。
ドパミン受容体はD1~D5に分類される。D3受容体およびD4受容体は思考の制御(統合失調症の陰性症状に限る)に関与しており,D2受容体の活性化は錐体外路系を制御する。しかしながら,受容体への結合親和性から機能的な応答(内因活性)を予測することはできない。例えば,ロピニロールはD3受容体に高い親和性を示すが,内因活性はD2受容体の活性化を介して生じる。
ノルアドレナリン
ノルアドレナリンは,ほとんどの交感神経節後線維と多くの中枢ニューロン(例,青斑および視床下部)の神経伝達物質である。前駆体であるチロシンがドパミンに変換された後,ドパミンβ-水酸化酵素により水酸化されてノルアドレナリンとなる。放出されて受容体と相互作用した後,ノルアドレナリンの一部はカテコールO-メチルトランスフェラーゼ(COMT)により分解され,残りは神経終末へ能動的に再取り込みされ,そこでMAOにより分解される。チロシン水酸化酵素,ドパミンβ-水酸化酵素,およびMAOがニューロン内のノルアドレナリン濃度を調節している。
アドレナリン受容体はα1(交感神経系のシナプス後膜),α2(交感神経系のシナプス前膜および脳のシナプス後膜),β1(心臓),β2(交感神経支配を受けるその他の構造)に分類される。
エンドルフィンおよびエンケファリン
エンドルフィンとエンケファリンはオピオイドである。
エンドルフィンは,多くの中枢ニューロン(例,視床下部,扁桃体,視床,青斑)を活性化する大きなポリペプチドである。細胞体には,α-,β-,γ-エンドルフィンの前駆体であるプロオピオメラノコルルチンと呼ばれる大きなポリペプチドが存在する。プロオピオメラノコルチンは軸索内を輸送されて複数の断片に開裂するが,その1つがβ-エンドルフィンであり,中脳水道周囲灰白質および大脳辺縁系に投射するニューロンと脳の主なカテコールアミンニューロンに認められる。放出されて受容体と相互作用した後,β-エンドルフィンはペプチダーゼによって加水分解される。
エンケファリンにはMet-エンケファリンとLeu-エンケファリンがあり,これらは多くの中枢ニューロン(例,淡蒼球,視床,尾状核,中心灰白質)に存在する小さなポリペプチドである。これらの前駆体であるプロエンケファリンが細胞体で産生され,これが特異的ペプチダーゼによって活性化ペプチドへと分断される。これらの物質は脊髄にも存在し,そこでは疼痛刺激を修飾する。脊髄後角における疼痛刺激の神経伝達物質は,グルタミン酸とサブスタンスPである。エンケファリンは,放出される神経伝達物質の量を減少させるとともに,シナプス後膜を過分極(より負にする)させ,活動電位の発生と中心後回レベルでの疼痛知覚を減弱させる。放出されてペプチド受容体と相互作用した後,エンケファリンは不活性の小さなペプチドおよびアミノ酸へと加水分解される。外因性のエンケファリンは急速に不活化されることから,これらの物質は臨床的には有用でない。代わりに,より安定な分子(例,モルヒネ)が鎮痛薬として使用されている。
エンドルフィン-エンケファリン(オピオイド)受容体は,μ1およびμ2(感覚運動統合および鎮痛に関与する),δ1およびδ2(運動の統合,認知機能,および鎮痛に関与),ならびにκ1,κ2,およびκ3(水分バランスの制御,鎮痛,および摂食行動に関与する)受容体に分類される。σ受容体は現在では非オピオイドとして分類されており,大半が海馬に局在し,PCPと結合する。新しいデータと薬理学的な推測に基づき,さらに多くの受容体サブタイプの存在が示唆されている。受容体タンパク質の合成過程で前駆体の構成要素に再構成が起きることで,受容体の変異体がいくつか生じる可能性がある(例,μオピオイド受容体には27のスプライス変異体がある)。2つの受容体が結合(二量体化)して新しい受容体を形成することもある。
その他の神経伝達物質
ダイノルフィンは,互いにアミノ酸配列の類似した7つのペプチドで構成されるグループである。これらはエンケファリンと同様にオピオイドである。
ペプチドであるサブスタンスPは,中枢ニューロン(手綱,黒質,基底核,延髄,および視床下部)に存在するほか,後根神経節に高濃度で存在する。その放出は,疼痛による強い求心性刺激によって誘発される。サブスタンスPは,疼痛および気分に対する神経学的応答を修飾し,また脳幹に局在するNK1A受容体の活性化を介して悪心および嘔吐を修飾する。
一酸化窒素(NO)は,ニューロンレベルの多くのプロセスに関与する不安定な気体である。NO合成酵素によってアルギニンから産生される。細胞内カルシウム濃度を上昇させる神経伝達物質(例,サブスタンスP,グルタミン酸,アセチルコリン)は,NO合成酵素を発現するニューロンにおいてNO合成を刺激する。NOは細胞内メッセンジャーである可能性があり,細胞外の二次ニューロンに拡散して生理反応を引き起こしたり(例,長期増強[特定のシナプス前およびシナプス後の応答を強化する―学習の一形態]),グルタミン酸の(NMDA受容体を介した)神経毒性(例,パーキンソン病,脳卒中,アルツハイマー病)を増強したりする可能性がある。
神経伝達における役割が十分に確立されていない物質としては,ヒスタミン,バソプレシン,血管作動性腸管ペプチド(VIP),カルノシン,ブラジキニン,コレシストキニン,ボンベシン,ソマトスタチン,コルチコトロピン放出因子,ニューロテンシンなどがあり,おそらくはアデノシンも該当する。
エンドカンナビノイドは,脳,内分泌系,および免疫系の機能を調節する内因性の脂質ベースの神経伝達物質である。
神経伝達の障害が関連する疾患
神経伝達物質の産生,放出,受容,分解,または再取り込みに変化を及ぼしたり,受容体の数や結合親和性を変化させたりする障害ないし物質は,神経または精神症状や疾患の原因となりうる(神経伝達の障害が関連する疾患の例の表を参照)。それらの疾患の多く(例,パーキンソン病,うつ病)は,神経伝達を修飾する薬剤によって軽快させることが可能である。