


Un neurone genera e propaga un potenziale d'azione lungo il proprio assone, quindi trasmette questo segnale attraverso una sinapsi rilasciando neurotrasmettitori, che innescano una risposta in un altro neurone o in una cellula effettrice (p. es., le cellule muscolari, la maggior parte delle cellule esocrine ed endocrine). I neurotrasmettitori consentono ai neuroni di comunicare tra loro. I neurotrasmettitori che vengono rilasciati si legano ai recettori in un altro neurone. I neuroni che rilasciano i neurotrasmettitori sono chiamati neuroni presinaptici. I neuroni che ricevono i segnali del neurotrasmettitore sono chiamati neuroni postsinaptici. In relazione al neurotrasmettitore e al recettore coinvolti, il segnale può stimolare o inibire la cellula che lo riceve. Altri fattori, tra cui farmaci e disturbi, influenzano la comunicazione tra i neuroni modulando la produzione e le azioni dei neurotrasmettitori tra cui
Il loro rilascio, la loro ricaptazione e la loro degradazione
Il numero e la funzione dei recettori dei neurotrasmettitori postsinaptici
A volte i segnali tra i neuroni avvengono nella direzione opposta (cosiddetta neurotrasmissione retrograda). In questi casi, i dendriti (i rami riceventi del neurone) sui neuroni postsinaptici rilasciano i neurotrasmettitori, che influenzano i recettori sui neuroni presinaptici. La trasmissione retrograda può inibire i neuroni presinaptici dal rilascio di neurotrasmettitori aggiuntivi, e può aiutare a controllare il livello di attività e di comunicazione tra i neuroni.
A livello del sistema nervoso centrale le interconnessioni sono complesse. Un impulso da un neurone all'altro può passare da
Assone al corpo cellulare
Assone a dendrite
Corpo cellulare a corpo cellulare
Dendrite a dendrite
Un neurone può ricevere simultaneamente molti stimoli, eccitatori e inibitori, da altri neuroni e integrare impulsi simultanei in vari pattern di scarica.
Propagazione
La propagazione del potenziale d'azione lungo un assone è di tipo elettrico ed è dovuta agli scambi di ioni sodio e potassio attraverso la membrana assonale. Un determinato neurone genera un potenziale d'azione identico dopo ogni stimolo, conducendolo a una velocità fissa lungo l'assone. La velocità dipende dal diametro assonale e dal grado di mielinizzazione e varia da 1-4 m/s nelle piccole fibre amieliniche fino a 75 m/s nelle fibre mieliniche di grosso calibro. La velocità di propagazione è maggiore nelle fibre mieliniche poiché la guaina mielinica è provvista di interruzioni ad intervalli regolari (nodi di Ranvier) in cui l'assone è scoperto. L'impulso elettrico si propaga da un nodo al successivo, saltando la porzione mielinizzata dell'assone. Pertanto, i processi patologici che alterano la guaina mielinica (p. es., la sclerosi multipla, sindrome di Guillain-Barré) interferiscono con la propagazione dell'impulso, causando diversi sintomi neurologici.
Trasmissione
La trasmissione dell'impulso è di natura chimica ed è causata dalla liberazione di specifici neurotrasmettitori dalla terminazione nervosa. I neurotrasmettitori si diffondono attraverso lo spazio sinaptico e si legano transitoriamente a recettori specifici sul neurone o sulla cellula effettrice adiacente. A seconda del recettore, la risposta può essere eccitatoria o inibitoria. Di solito, i neuroni non sono in contatto diretto ma comunicano tramite la trasmissione di neurotrasmettitori attraverso le sinapsi. In alcune circostanze, i neuroni vicini possono comunicare utilizzando impulsi elettrici attraverso una giunzione gap.
Il corpo della cellula nervosa produce enzimi che sintetizzano la maggior parte dei neurotrasmettitori, i quali vengono immagazzinati in vescicole a livello della terminazione sinaptica (vedi figura Neurotrasmissione). La quantità immagazzinata in una singola vescicola (in genere diverse migliaia di molecole) costituisce un quanto. Un potenziale d'azione di membrana che raggiunge il terminale sinaptico determina l'apertura dei canali del calcio dell'assone; l'ingresso del calcio libera molecole di neurotrasmettitore da varie vescicole, mediante la fusione delle membrane delle vescicole con la membrana della terminazione nervosa. La fusione delle membrane genera un'apertura attraverso cui le molecole vengono rilasciate nello spazio sinaptico per esocitosi.
Un tipo di sinapsi, la sinapsi elettrica, non utilizza i neurotrasmettitori; i canali ionici collegano direttamente il citoplasma dei neuroni presinaptici a quello dei neuroni postsinaptici. Questo tipo di trasmissione è il più veloce.
Segnali eccitatori ed inibitori
La reazione innescata dal rilascio del neurotrasmettitore può eccitare o attivare il neurone postsinaptico o inibire o bloccare la sua attività. I neuroni postsinaptici ricevono più segnali neurotrasmettitori e segnali elettrici da molti neuroni. Il neurone ricevente alla fine somma assieme gli stimoli e, se vengono ricevuti più segnali eccitatori, il neurone si attiva e invia i segnali ad altri neuroni. Se la somma dei segnali è inibitoria, il neurone non si attiva e non influenza l'attività di altri neuroni. Questa somma di risposte è chiamata sommatoria. I neurotrasmettitori facilitano così una rapida comunicazione tra i neuroni modificando l'attivazione del potenziale d'azione.
Altre forme di sommatoria comprendono
Sommatoria spaziale: quando si ricevono più impulsi in diverse posizioni del neurone e il neurone li sintetizza
Sommatoria temporale: quando gli impulsi si ricevono in un breve periodo di tempo e poi si sommano
Affinché un neurone generi un segnale e si attivi, deve raggiungere un potenziale di soglia. Il potenziale di soglia è prodotto da un aumento netto dell'afflusso di sodio nella cellula durante lo scambio di ioni di sodio e di potassio. Quando una quantità sufficiente di sodio entra nella cellula, si raggiunge la soglia; quando si raggiunge la soglia, si attiva un potenziale d'azione; esso viaggia lungo la membrana del neurone. La soglia deve essere raggiunta per generare un potenziale d'azione.
Neurotrasmissione
I potenziali d'azione aprono i canali del calcio assonali (non mostrato). Il Ca++ innesca il rilascio dei neurotrasmettitori dalle vescicole in cui sono immagazzinati. Le molecole dei neurotrasmettitori riempiono lo spazio sinaptico. Alcune si legano ai recettori postsinaptici, generando una risposta. Le altre sono riassorbite nell'assone e immagazzinate oppure si diffondono nei tessuti circostanti. |

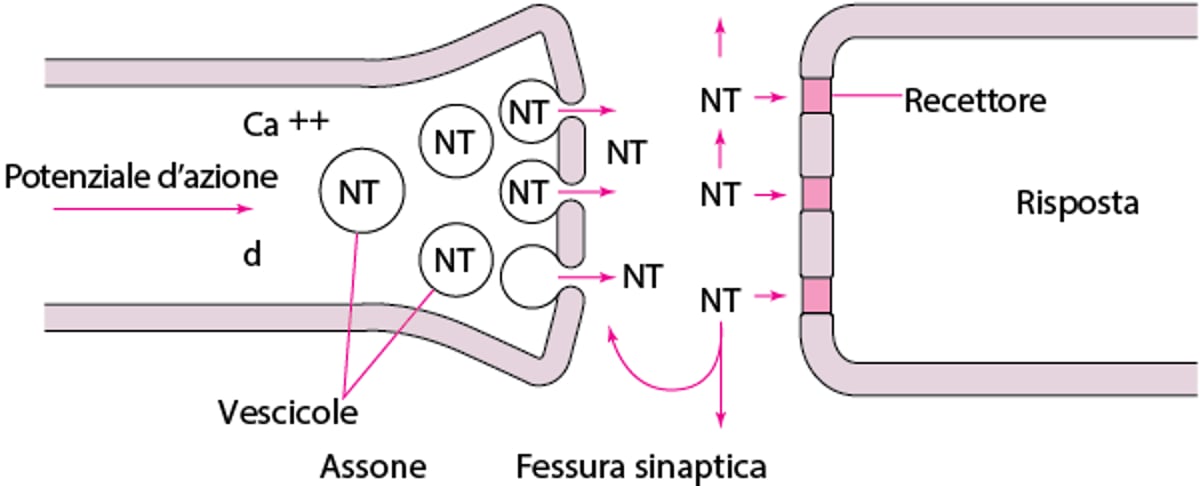
La quantità di neurotrasmettitore nella terminazione sinaptica è tipicamente indipendente dall'attività nervosa ed è mantenuta relativamente costante dalla modifica della ricaptazione dei precursori del neurotrasmettitore o dell'attività degli enzimi coinvolti nella sintesi o nella degradazione del neurotrasmettitore. La stimolazione dei recettori presinaptici può ridurre la sintesi presinaptica del neurotrasmettitore, mentre il loro blocco può aumentarla.
L'interazione neurotrasmettitore-recettore deve essere presto interrotta per porre fine all'azione continua del neurotrasmettitore e/o consentire una rapida e ripetuta attivazione dei recettori. I neurotrasmettitori che hanno interagito con i recettori possono andare incontro a una delle seguenti condizioni:
Essi possono essere rapidamente riassorbiti nelle terminazioni nervose presinaptiche mediante processi attivi adenosina trifosfato (ATP)-dipendenti (reuptake) per il ricircolo o la distruzione.
Essi possono essere eliminati da enzimi in prossimità dei recettori.
Essi possono diffondersi nella zona circostante ed essere rimossi.
I neurotrasmettitori ricaptati dalle terminazioni nervose vengono nuovamente immagazzinati in granuli o vescicole nell'assone terminale per poter essere riutilizzati.
Il malfunzionamento di questi processi può causare malattie cliniche. Per esempio, si ipotizza che la perdita di memoria nella malattia di Alzheimer coinvolga l'insufficienza del neurotrasmettitore acetilcolina nelle sinapsi, che media la deposizione di nuovi ricordi. Alcuni farmaci (p. es., donepezil, galantamina, rivastigmina) bloccano l'enzima acetilcolinesterasi (che scompone l'acetilcolina) e quindi aumentano la quantità di acetilcolina nella sinapsi. Come risultato, il funzionamento della memoria può migliorare.
Alcuni tipi di neuroni singoli possono rilasciare due o più neurotrasmettitori diversi (cosiddetta cotrasmissione); per esempio, l'acetilcolina e il glutammato. Più neurotrasmettitori possono agire su un singolo neurone postsinaptico o colpire più neuroni postsinaptici. La cotrasmissione consente la complessa comunicazione tra i neuroni per controllare diversi eventi nel sistema nervoso centrale e nel sistema nervoso periferico.
I neurotrasmettitori possono anche facilitare cambiamenti più a lungo termine che coinvolgono percorsi aggiuntivi come i cambiamenti nell'attività dei geni e delle proteine.
Recettori
I recettori per i neurotrasmettitori sono complessi proteici disposti lungo la membrana cellulare. La loro natura determina la funzione eccitatoria o inibitoria di un certo neurotrasmettitore. I recettori che vengono stimolati continuamente dai neurotrasmettitori o dai farmaci vanno incontro a desensibilizzazione (down-regulation); quelli che non vengono stimolati dal relativo neurotrasmettitore o che vengono bloccati cronicamente dai farmaci diventano ipersensibili (up-regulation). La up- o down-regulation dei recettori condiziona fortemente lo sviluppo della tolleranza e della dipendenza fisica. Questi concetti sono particolarmente importanti nei trapianti d'organo o tessuto, in cui la denervazione priva i recettori del proprio neurotrasmettitore; di conseguenza, gli organi trapiantati possono diventare eccessivamente sensibili alla stimolazione neurale. I sintomi di astinenza possono essere spiegati almeno in parte con un meccanismo di rimbalzo dovuto a un'alterazione dell'affinità o della densità recettoriali.
La maggior parte dei neurotrasmettitori interagisce principalmente con i recettori postsinaptici, ma alcuni recettori sono localizzati sui neuroni presinaptici, consentendo un fine controllo del rilascio del neurotrasmettitore.
Una famiglia di recettori, chiamati recettori ionotropici (p. es., recettori di N-metil-d-glutammato, kinate-quisqualato, nicotinico, glicina e acido gamma-aminobutirrico [GABA]), è costituita da canali ionici che si aprono quando si legano al neurotrasmettitore promuovendo una risposta molto rapida. Nell'altra famiglia, quella dei recettori metabotropici (p. es., la serotonina, i recettori alfa- e beta-adrenergici e dopaminergici), i neurotrasmettitori interagiscono con le proteine G e attivano un'altra molecola (il secondo messaggero come l'adenosina-monofosfato ciclico) che catalizza una catena di eventi attraverso la mobilizzazione del calcio o la fosforilazione proteica, o entrambe; i cambiamenti cellulari mediati dal secondo messaggero sono più lenti e consentono una più fine regolazione della risposta rapida del neurotrasmettitore ionotropico. Inoltre, molti neurotrasmettitori attivano più recettori specifici rispetto ai secondi messaggeri.
Principali neurotrasmettitori e recettori
Almeno 100 sostanze possono agire come neurotrasmettitori; circa 18 sono di primaria importanza. Molte sono presenti in forme che differiscono lievemente tra loro. I neurotrasmettitori possono essere raggruppati in diverse classi, come
Piccole molecole (p. es., glutammato, acido gamma-aminobutirrico, glicina, adenosina, acetilcolina, serotonina, istamina, noradrenalina)
Neuropeptidi (p. es., endorfine)
Molecole gassose (p. es., ossido nitrico, monossido di carbonio)
Endocannabinoidi
Glutammato e aspartato
Questi aminoacidi (glutammato e aspartato) sono i principali neurotrasmettitori eccitatori nel sistema nervoso centrale. Sono presenti nella corteccia, nel cervelletto e nel midollo spinale. Nei neuroni, la sintesi dell'ossido nitrico aumenta in risposta al glutammato. Un eccesso di glutammato può essere tossico, determinando un aumento del calcio intracellulare, dei radicali liberi e dell'attività proteasica. Questi neurotrasmettitori possono contribuire alla tolleranza alla terapia con oppiacei e mediare l'iperalgesia.
I recettori glutammatergici (stimolati dal glutammato e meno fortemente dall'aspartato) sono classificati come recettori N-metil-d-aspartato e non-N-metil-d-aspartato. La fenciclidina (anche nota come polvere d'angelo) e la memantina (utilizzata per il trattamento della malattia di Alzheimer) si legano ai recettori N-metil-d-aspartato.
Acido gamma-aminobutirrico
L'acido gamma-aminobutirrico (GABA, gamma-aminobutyric acid) è il principale neurotrasmettitore inibitorio nel cervello. È un aminoacido che deriva dal glutammato, il quale viene decarbossilato dalla glutammato decarbossilasi. Dopo l'interazione con i propri recettori, l'acido gamma-aminobutirrico viene attivamente riassorbito nei terminali nervosi e metabolizzato. La glicina, che presenta un'azione simile all'acido gamma-aminobutirrico, si trova principalmente negli interneuroni (cellule di Renshaw) del midollo spinale e nei circuiti che determinano il rilassamento dei muscoli antagonisti.
I recettori dell'acido gamma-aminobutirrico sono classificati come GABA-A (che attivano i canali del cloro) e GABA-B (che promuovono la formazione di adenosina-monofosfato ciclico). I recettori GABA-A rappresentano il sito d'azione di molti farmaci neuroattivi, incluse le benzodiazepine, i barbiturici, la picrotossina e il muscimolo. L'alcol si lega anche ai recettori GABA-A. I recettori GABA-B sono attivati dal baclofen, utilizzato per il trattamento della spasticità muscolare (p. es., nella sclerosi multipla).
Serotonina
La serotonina (5-idrossitriptamina, 5-HT) è generata dai nuclei del rafe e dai neuroni della linea mediana del ponte e della porzione superiore del tronco encefalico. Il triptofano viene idrossilato dalla triptofano idrossilasi in 5-idrossitriptofano e quindi decarbossilato a serotonina. I livelli di serotonina sono regolati dalla captazione del triptofano e dalla monoamino ossidasi intraneuronale, che degrada la serotonina. In ultima analisi, la serotonina viene escreta nelle urine come acido 5-idrossiindolacetico o 5-HIAA.
I recettori serotoninergici (5-idrossitriptamina, presenti in almeno 15 sottotipi diversi) sono classificati come 5-HT1 (con 4 sottotipi), 5-HT2 e 5-HT3. Gli agonisti selettivi dei recettori della serotonina (p. es., sumatriptan) possono bloccare gli attacchi emicranici. Gli inibitori selettivi della ricaptazione della serotonina possono anche essere usati per trattare diversi disturbi di salute mentale (p. es., depressione, ansia, disturbo ossessivo-compulsivo, disturbo da stress post-traumatico).
Acetilcolina
L'acetilcolina rappresenta il principale neurotrasmettitore dei motoneuroni bulbospinali, delle fibre autonomiche pregangliari, delle fibre colinergiche postgangliari (parasimpatiche) e di molti neuroni del sistema nervoso centrale (p. es., gangli della base, corteccia motoria). Viene sintetizzata a partire dalla colina e dall'acetil-coenzima A a opera della colina acetiltransferasi e la sua azione viene rapidamente interrotta per mezzo di un'idrolisi locale in colina e acetato a opera dell'acetilcolinesterasi. I livelli di acetilcolina sono regolati dalla colina acetiltransferasi e dalla captazione della colina. I livelli di questo neurotrasmettitore sono ridotti nei pazienti affetti dalla malattia di Alzheimer.
I recettori colinergici sono classificati come nicotinici N1 (nel muscolo scheletrico e nella giunzione neuromuscolare) o N2 (nel sistema nervoso centrale e periferico, che include il sistema nervoso parasimpatico e simpatico e la midollare del surrene) o muscarinici da M1 a M5 (ampiamente distribuito nel sistema nervoso centrale). L'M1 è presente nel sistema nervoso autonomo, nello striato, nella corteccia e nell'ippocampo; l'M2 è presente nel sistema nervoso autonomo, nel cuore, nella muscolatura liscia intestinale, nel rombencefalo e nel cervelletto.
Dopamina
La dopamina interagisce con i recettori di alcune fibre dei nervi periferici e di molti neuroni centrali (p. es., nella substantia nigra, nel mesencefalo, nell'area tegmentale ventrale e nell'ipotalamo). L'aminoacido tirosina viene incorporato dai neuroni dopaminergici e convertito dalla tirosino-idrossilasi in 3,4-diidrossifenilalanina, la quale viene decarbossilata dalla decarbossilasi degli l-aminoacidi aromatici in dopamina. Dopo il rilascio e l'interazione con i recettori, la dopamina viene recuperata in maniera attiva (ricaptazione) dal terminale nervoso. La tirosina-idrossilasi e le monoamino ossidasi (che degradano la dopamina) regolano i livelli di dopamina nei terminali nervosi.
I recettori dopaminergici sono classificati da D1 a D5. I recettori D3 e D4 hanno un ruolo nel controllo della produzione del pensiero (limitando i sintomi negativi della schizofrenia); l'attivazione del recettore D2 controlla il sistema extrapiramidale. Tuttavia, l'affinità recettoriale non è in grado di prevedere la risposta funzionale (attività intrinseca). Per esempio, il ropinirolo, che ha alta affinità per il recettore D3, ha attività intrinseca mediante l'attivazione di recettori D2.
Noradrenalina
La noradrenalina è il neurotrasmettitore della maggior parte delle fibre simpatiche postgangliari e di molti neuroni centrali (p. es., nel locus coeruleus e nell'ipotalamo). Il precursore tirosina è convertito in dopamina, la quale viene idrossilata dalla dopamina beta-idrossilasi in noradrenalina. Dopo il rilascio e l'interazione con i recettori, parte della noradrenalina viene degradata dalla catecolo O-metiltrasferasi, mentre la parte restante viene attivamente riassorbita nel terminale nervoso, dove viene degradata dalle monoamino ossidasi. La tirosina-idrossilasi, la dopamina beta-idrossilasi e le monoamino ossidasi regolano i livelli intraneuronali della noradrenalina.
I recettori adrenergici sono classificati come alfa-1 (postsinaptici nel sistema simpatico), alfa-2 (presinaptici nel sistema simpatico e postsinaptici nell'encefalo), beta-1 (nel cuore) o beta-2 (in altre strutture ad innervazione simpatica).
Endorfine ed encefaline
Endorfine ed encefaline sono oppiacei.
Le endorfine sono grandi polipeptidi che attivano un gran numero di neuroni centrali (p. es., a livello dell'ipotalamo, dell'amigdala, del talamo e del locus coeruleus). I corpi cellulari contengono un grande polipeptide denominato pro-opiomelanocortina, il precursore delle alfa-, beta- e gamma-endorfine. La pro-opiomelanocortina è trasportata lungo l'assone e scissa in frammenti; uno costituisce la beta-endorfina, contenuta nei neuroni che proiettano alla sostanza grigia periacqueduttale, alle strutture limbiche e ai principali neuroni cerebrali contenenti catecolamine. Dopo il rilascio e l'interazione con i recettori, la beta-endorfina viene idrolizzata dalle peptidasi.
Le encefaline comprendono la met-encefalina e la leu-encefalina, che sono piccoli polipeptidi presenti in molti neuroni centrali (p. es., nel globo pallido, nel talamo, nel caudato e nella sostanza grigia centrale). Il loro precursore, la proencefalina, viene prodotto nel corpo cellulare e quindi scisso da peptidasi specifiche nei peptidi attivi. Tali molecole sono localizzate anche nel midollo spinale, dove agiscono come modulatori degli stimoli dolorifici. I neurotrasmettitori degli stimoli dolorifici nel corno posteriore del midollo spinale sono il glutammato e la sostanza P. Le encefaline riducono la quantità di neurotrasmettitore rilasciato e iperpolarizzano (rendono più negativa) la membrana postsinaptica, riducendo la generazione dei potenziali d'azione e la percezione dolorosa a livello del giro postcentrale. Dopo il rilascio e l'interazione con i recettori peptidergici, le encefaline vengono idrolizzate in peptidi inattivi più piccoli e in aminoacidi. La rapida inattivazione delle encefaline esogene previene la possibilità che possano essere utili dal punto di vista clinico. Molecole più stabili (p. es., la morfina) vengono infatti utilizzate come analgesici.
I recettori per endorfina-encefalina (oppiacei) sono classificati come mu-1 e mu-2 (che interessano l'integrazione sensomotoria e l'analgesia), delta-1 e delta-2 (che interessano l'integrazione motoria, le funzioni cognitive e l'analgesia) e kappa-1, kappa-2 e kappa-3 (che interessano la regolazione dell'equilibrio idrico, l'analgesia e l'assunzione di cibo). I recettori sigma, attualmente classificati come non oppiacei e localizzati principalmente a livello dell'ippocampo, legano la fenciclidina. Dati recenti suggeriscono l'esistenza di molti altri sottotipi recettoriali, con implicazioni farmacologiche. Componenti del precursore molecolare della proteina recettoriale possono andare incontro a riarrangiamento durante la sintesi del recettore e produrre diverse varianti recettoriali (p. es., 27 varianti di splicing del recettore mu degli oppiacei). Inoltre, 2 recettori possono combinarsi (dimerizzare) in modo da formare un nuovo recettore.
Altri neurotrasmettitori
Le dinorfine sono un gruppo di 7 peptidi con sequenze aminoacidiche simili. Esse, al pari delle encefaline, sono oppiacei.
La sostanza P, un peptide, si trova nei neuroni centrali (abenula, substantia nigra, gangli della base, bulbo e ipotalamo) ed è estremamente concentrata nei gangli delle radici dorsali. Il suo rilascio è innescato da intensi stimoli dolorosi afferenti. Essa modula la risposta neurale al dolore e l'umore; modula la nausea e il vomito attraverso l'attivazione di recettori NK1A che sono localizzati nel tronco encefalico.
L'ossido nitrico è un gas instabile che media numerosi processi neuronali. Viene generato dall'arginina attraverso la NO-sintetasi. I neurotrasmettitori che determinano un aumento intracellulare di calcio (p. es., la sostanza P, il glutammato, l'acetilcolina) stimolano la sintesi di NO nei neuroni che esprimono la NO-sintetasi. L'ossido nitrico può comportarsi da messaggero intracellulare; esso può diffondersi da una cellula in un secondo neurone e generare risposte fisiologiche (p. es., il potenziamento a lungo termine [rafforzamento di talune risposte presinaptiche e postsinaptiche, una forma di apprendimento]) o incrementare la neurotossicità del glutammato (N-metil-d-aspartato recettore-mediato, p. es., nel morbo di Parkinson, nell'ictus o nella malattia di Alzheimer). Il monossido di azoto colpisce altri neurotrasmettitori (p. es., acido gamma-aminobutirrico e acetilcolina) modificando l'afflusso di calcio nelle cellule per aumentare il rilascio di altri neurotrasmettitori.
Ulteriori neurotrasmettitori gassosi comprendono il monossido di carbonio (CO) e l'acido solfidrico (H2S). Questi trasmettitori sono prodotti in cellule di tutto il corpo (compreso il cervello). La CO endogena è generata dal metabolismo dell'eme e può partecipare a processi che coinvolgono la generazione di febbre, l'infiammazione, la sopravvivenza cellulare e il controllo della dilatazione dei vasi sanguigni. Diversi enzimi sono coinvolti nella produzione di H2S, che si ritiene sia necessario per il cervello per formare e conservare i ricordi.
Sostanze con un ruolo meno definito nella neurotrasmissione comprendono l'istamina, la vasopressina, il peptide vasoattivo intestinale, la carnosina, la bradichinina, la colecistochinina, la bombesina, la somatostatina, il fattore di rilascio della corticotropina, la neurotensina e, probabilmente, l'adenosina.
Gli endocannabinoidi sono neurotrasmettitori endogeni a base lipidica che modulano le funzioni cerebrali, endocrine e del sistema immunitario.
Patologie associate a difetti della neurotrasmissione
I processi patologici o le sostanze che alterano la produzione, il rilascio, la ricezione, l'eliminazione o la ricaptazione dei neurotrasmettitori o che modificano il numero e l'affinità dei recettori possono causare sintomi e malattie neurologiche o psichiatriche (vedi tabella Esempi di patologie associate a difetti della neurotrasmissione). I farmaci che modificano la neurotrasmissione possono alleviare molti di questi disturbi (p. es., morbo di Parkinson, depressione).







