


Der Übergang vom Leben in utero zum Leben außerhalb des Mutterleibes bringt viele Veränderungen der Physiologie und Funktionen mit sich. Siehe auch Perinatale Probleme.
Neonataler Bilirubin-Metabolismus
(Siehe auch Leberstruktur-und Funktion und Neonatale Hyperbilirubinämie.)
Alte oder geschädigte Erythrozyten werden durch das retikuloendotheliale System, das Häm zu Bilirubin abbaut (1 g Hämoglobin ergibt 35 mg Bilirubin), aus dem Kreislauf entfernt. Dieses Bilirubin bindet sich an Albumin und wird zur Leber transportiert, wo es in die Hepatozyten übertragen wird. Das Bilirubin wird dort durch die Glukuronyltransferase mit Uridindiphosphoglukuronsäure (UDPGA) zu Bilirubindiglukuronid (konjugiertes Bilirubin) umgewandelt und aktiv in die Gallengänge sezerniert.
Von dort gelangt das Bilirubindiglukuronid im Gastrointestinaltrakt in das Mekonium, kann jedoch nicht ausgeschieden werden, da der Fetus normalerweise keine Stühle absetzt. Das Enzym Beta-Glucuronidase, das in der Dünndarmluminalbürste des Fötus vorhanden ist, wird in das Darmlumen freigesetzt, wo es Bilirubinglukuronid dekonjugiert; freies (unkonjugiertes) Bilirubin wird dann aus dem Darmtrakt resorbiert und gelangt wieder in den fetalen Kreislauf.
Freies (unkonjugiertes) Bilirubin wird dann wieder aus dem Intestinaltrakt resorbiert und in den fetalen Kreislauf aufgenommen. Das fetale Bilirubin wird schließlich ausgeschieden, in dem es die Plazenta entlang eines Konzentrationsgradienten passiert und in das mütterliche Plasma gelangt und schließlich von der mütterlichen Leber konjugiert und ausgeschieden wird.
Unmittelbar nach der Geburt wird die Verbindung mit der Plazenta beendet. Zwar kann die Leber des Neugeborenen das anfallende Bilirubin weiter aufnehmen konjugieren und in die Galle und von dort in den Stuhl ausscheiden. Da aber die physiologische intestinale Bakterienbesiedlung beim Neugeborenen noch nicht ausgebildet ist, kann das Bilirubin im Darm nicht zu Urobilinogen oxidiert werden. Das unveränderte Bilirubin verbleibt im Stuhl, was diesem die typische hellgelbe Farbe verleiht. Ebenso wie beim Fetus enthält der Gastrointestinaltrakt des Neugeborenen Beta-Glukuronidase, so dass ein Teil des Bilirubins dekonjugiert wird.
Fütterungen führen zum gastrokolischen Reflex, sodass Bilirubin im Stuhl ausgeschieden wird, bevor es dekonjugiert und reabsorbiert werden kann. Dagegen wird bei vielen Neugeborenen das unkonjugierte Bilirubin vom Darmlumen reabsorbiert und wieder dem Kreislauf zugeführt (enterohepatischer Kreislauf des Bilirubins), was dann zur physiologischen Hyperbilirubinämie und zum Ikterus beiträgt. Der erhöhte Hämatokrit und die kürzere Lebensdauer der Erythrozyten bei Neugeborenen führen zu einer erhöhten Bilirubinproduktion und tragen ebenfalls zur physiologischen Hyperbilirubinämie bei.
Neonataler Herz-Kreislauf-Funktion
Im fetalen Kreislauf fließt das Blut mit Rechts-Links-Shunts durch den offenen Ductus arteriosus Botalli (der die Pulmonalarterie und die Aorta verbindet) und das Foramen ovale (das den rechten und linken Vorhof verbindet) von rechts nach links, vorbei an der nichtoxygenierten Lunge. Dieser sog. Rechts-Links-Shunt beruht im Wesentlichen auf dem hohen pulmonalarteriellen und dem relativ niedrigen systemischen (wozu auch die Plazenta zu zählen ist) Widerstand. Ungefähr 90–95% des vom rechten Ventrikel ausgeworfenen Blutes fließen direkt in die systemische Zirkulation und somit an den Lungen vorbei.
Durch den niedrigen fetalen PaO2 (ca. 25 mmHg) und durch lokal produzierte Prostaglandine wird der Ductus arteriosus Botalli offen gehalten. Das Foramen ovale wird durch unterschiedliche Druckverhältnisse in den Vorhöfen offen gehalten: der linksatriale Druck ist aufgrund des geringen Blutrückflusses aus der Lunge relativ niedrig, der rechtsatriale Druck hingegen aufgrund des großen Blutrückflusses aus der Plazenta vergleichsweise hoch.
Normale Zirkulation bei einem Fötus
Im Fötus wurde Blut, das in die rechte Seite des Herzens eintritt, bereits über die Plazenta mit Sauerstoff angereichert. Da die Lunge nicht belüftet ist, muss nur eine geringe Menge Blut durch die Lungenarterie fließen. Das meiste Blut von der rechten Seite des Herzens umgeht die Lunge durch die.
Normalerweise schließen diese beiden Strukturen kurz nach der Geburt. |

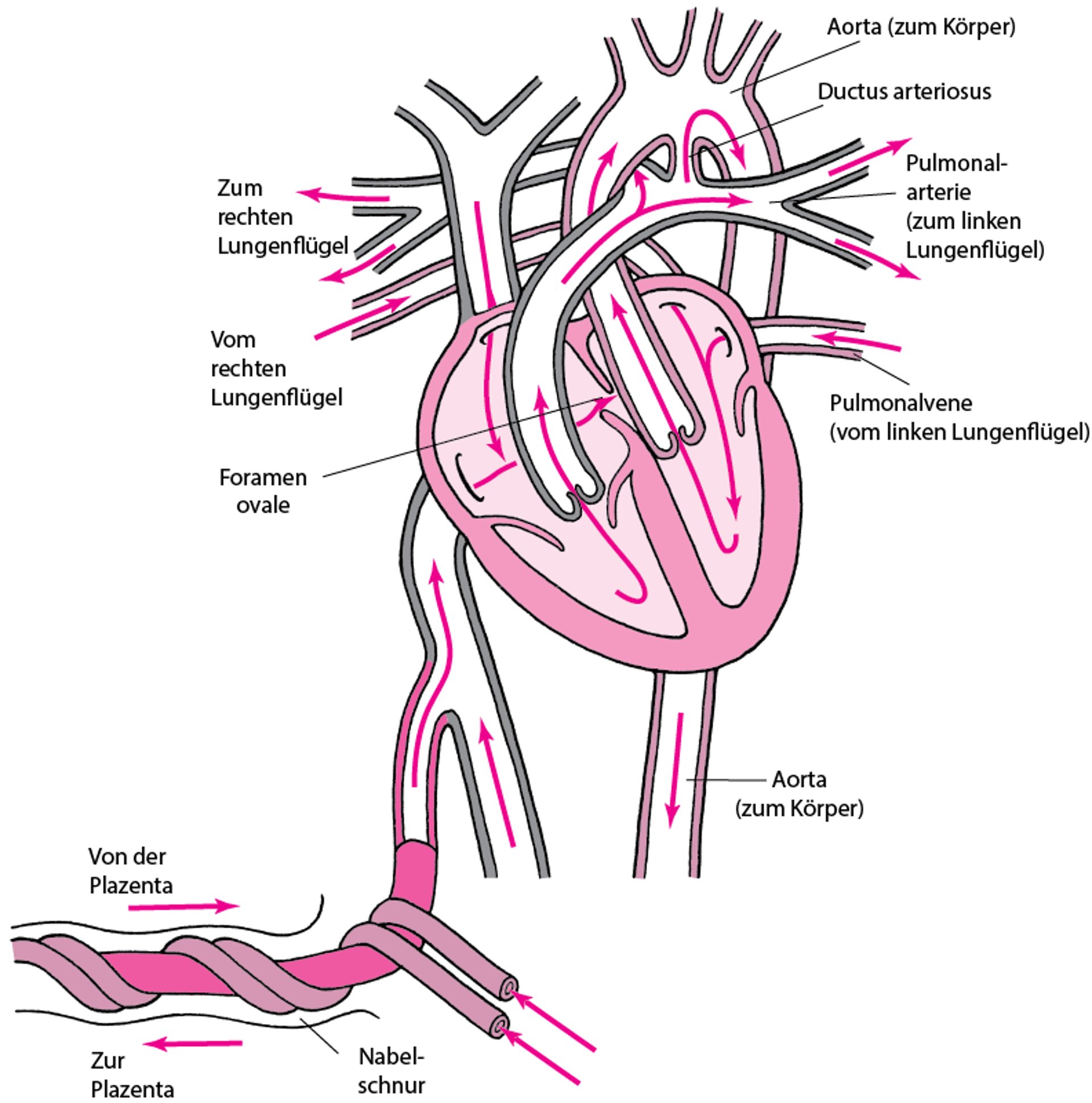
Nach dem ersten Atemzug kommt es zu grundlegenden Veränderungen in diesem System, was zu einem vermehrten pulmonalen Blutfluss und zum funktionalen Verschluss des Foramen ovale führt. Als Folge der Vasodilatation durch die Ausdehnung der Lunge, des angestiegenen PaO2 und des verminderten PaCO2 sinkt der pulmonalarterielle Widerstand akut ab. Zudem vermindert sich durch die elastischen Kräfte der Rippen und des Thorax der interstitielle pulmonale Druck, was zu einer weiteren Erhöhung des Blutflusses durch die Lungenkapillaren führt. Erhöhter venöser Rückstrom aus der Lunge erhöht den linken atrialen Druck, wodurch die Druckdifferenz zwischen dem linken und rechten Vorhof reduziert wird; Dieser Effekt trägt zur funktionalen Schließung des Foramen ovale bei.
Sobald sich ein normaler Blutfluss in der Lunge eingestellt hat, erhöht sich der venöse Rückstrom von den Lungen, woraufhin der linksatriale Druck ansteigt. Durch die Atmung wird der PaO2 erhöht, was zu einer Konstriktion der Nabelarterien führt. Der plazentare Blutfluss wird vermindert oder unterbrochen, was zu einem verminderten Blutfluss in den rechten Vorhof führt. Somit verringert sich der Druck im rechten Vorhof, während der linksatriale Druck steigt; als Ergebnis werden die beiden Komponenten des fetalen interatrialen Septum (Septum primum und Septum secundum) zusammengeschoben, was zur Durchflussunterbrechung durch das Foramen ovale führt. Bei den meisten Personen verschmelzen die zwei Septen schließlich und das Foramen ovale existiert nicht mehr.
Kurz nach der Geburt übersteigt der systemische den pulmonalarteriellen Widerstand; die fetalen Kreislaufverhältnisse haben sich umgekehrt. Dementsprechend kommt es zu einer Umkehr des Blutflusses durch den Ductus arteriosus und somit zu einem Links-Rechts-Shunt (sog. Übergangskreislauf). Dieser Zustand dauert von kurz nach der Geburt (wenn der pulmonale Blutfluss zunimmt und es zu einem funktionellen Verschluss des Foramen ovale kommt) bis zu 24–72 Stunden, wenn sich der Ductus arteriosus verengt. Das Blut, das den Ductus arteriosus und die Vasa vasorum von der Aorta aus durchströmt, hat einen hohen PO2, was zusammen mit Veränderungen im Prostaglandinmetabolismus zu einer Konstriktion und zum Verschluss des Ductus arteriosus führt. Sobald dieser verschlossen ist, bestehen Kreislaufverhältnisse wie beim Erwachsenen. Die beiden Ventrikel pumpen nun seriell und es bestehen keine wesentlichen Shunts mehr zwischen dem pulmonalen und systemischen Kreislaufsystem.
Pränataler Stress, postnataler Stress und anatomische Unterschiede, die zum Fortbestehen eines erhöhten pulmonalen Gefäßwiderstands nach der Geburt führen, können zu einer persistierenden pulmonalen Hypertonie des Neugeborenen führen, die früher als persistierende fetale Zirkulation bezeichnet wurde. Hypoxämie und Azidose verursachen eine Verengung der pulmonalen Arteriolen und eine Erweiterung des Ductus arteriosus, was die zuvor beschriebenen Prozesse umkehrt und zu einem Rechts-Links-Shunt durch den Ductus arteriosus, das Foramen ovale oder beides führt.
Ziel der Behandlung ist es, die Bedingungen, die die pulmonale Vasokonstriktion verursacht haben, rückgängig zu machen und die zugrunde liegenden Erkrankungen zu behandeln.
Neonatale endokrine Funktion
(Siehe auch Übersicht über das endokrine System.)
Der Fetus ist vollständig von der transplazentaren Glukoseversorgung durch die Mutter abhängig und trägt nicht zur Glukoseproduktion bei. Bereits früh in der Schwangerschaft beginnt der Fetus, hepatische Glykogenspeicher anzulegen, wobei der größte Anteil in der 2. Hälfte des 3. Hälfte des 3. Die Glukoseversorgung des Neugeborenen endet mit der Durchtrennung der Nabelschnur, gleichzeitig steigen die Spiegel von zirkulierendem Adrenalin, Noradrenalin und Glukagon, während die Insulinspiegel absinken. Durch diese Veränderungen werden die Gluconeogenese und die Mobilisation der Glykogenspeicher in der Leber stimuliert.
Bei gesunden Neugeborenen wird der tiefste Glukosespiegel etwa 30–90 Minuten nach der Geburt erreicht; danach ist normalerweise eine Glukosehomöostase gewährleistet. Ein sehr hohes Risiko für eine neonatale Hypoglykämie haben Kinder mit verminderten Glykogenspeichern (SGA-Kinder und frühgeborene Kinder), schwerkranke Neugeborene mit vermehrtem Glukosekatabolismus und Kinder diabetischer Mütter (sekundärer fetaler Hyperinsulinismus).
Beim Fetus ist die Plazenta die Hauptquelle für Mineralien, die für die Knochenmineralisierung notwendig sind, und zwar durch den aktiven Transport von Kalzium, Magnesium und Phosphor, der durch mütterliches und plazentares Parathormon (PTH) und PTH-verwandtes Protein (PTHrP) stimuliert wird. Beim Neugeborenen kontrollieren die Nieren und der Darm die Abgabe und Ausscheidung dieser Mineralien. Die fetale PTH-Produktion wird durch hohe zirkulierende Kalziumspiegel unterdrückt, die von der Plazenta geliefert werden. Wenn die Nabelschnur bei der Geburt durchtrennt wird, hört die plazentare Infusion von Kalzium auf, was zu einem 20- bis 30%igen Abfall des Serumkalziumspiegels führt, der nach 24–48 Stunden einen Tiefpunkt erreicht. Gleichzeitig steigt der Serumphosphor an. Es gibt eine Verzögerung, bevor die Nebenschilddrüsen bei Neugeborenen voll ansprechbar sind (physiologischer Hypoparathyreoidismus) und das Serum-PTH als Reaktion auf den Rückgang des Serumkalziums ansteigt. Die Kalzium- und Phosphorwerte stabilisieren sich dann auf dem Niveau der Neugeborenen, und die Mineralienhomöostase des Skeletts wird von Darm und Nieren kontrolliert.
Literatur zur endokrinen Funktion bei Neugeborenen
1. Kovacs CS: Calcium, phosphorus, and bone metabolism in the fetus and newborn. Early Hum Dev 91(11):623–628, 2015. doi: 10.1016/j.earlhumdev.2015.08.007
Neonatale hämopoetische Funktion
(Siehe auch Perinatale Anämie.)
Die fetale Erythropoese findet im Dottersack zwischen 2 und 10 Schwangerschaftswochen statt, in der Leber, dem Hauptort der Erythropoese, bis zur 18. Schwangerschaftswoche und im Knochenmark ab etwa der 18. Schwangerschaftswoche und wird zum primären Ort bis zur 30. Schwangerschaftswoche. Eine kutane extramedulläre Hämatopoese kann bei Neugeborenen beobachtet werden, die an einer schweren Anämie leiden (z. B. kongenitale Infektion, hämolytische Anämie) (1).
Die Produktion von Erythrozyten wird in utero ausschließlich durch fetales Erythropoetin reguliert, mütterliches Erythropoetin kann die Plazenta nicht passieren. Fetalzellen enthalten etwa 55 bis 90% des fetalen Hämoglobins (Hämoglobin F oder HbF), das eine hohe Sauerstoffaffinität aufweist. Dadurch wird ein hoher Sauerstoff-Konzentrationsgradient in der Plazenta aufrecht erhalten und somit ein reichlicher Sauerstofftransfer vom mütterlichen zum kindlichen Kreislauf gewährleistet. Nach der Geburt ist diese hohe Sauerstoffaffinität nicht mehr so nützlich, da der Sauerstoff weniger leicht an die Gewebe abgegeben wird. Dies wirkt sich besonders schädlich bei schwerwiegenden pulmonalen oder kardialen Krankheiten mit Hypoxie aus.
Der Wechsel von fetalem zu adultem Hämoglobin beginnt bereits vor der Geburt; durch einen bisher nicht näher bekannten Mechanismus ändert sich der Ort der Erythropoetinproduktion von der Leber zu den sensitiveren peritubulären Zellen der Niere. Durch den abrupten Anstieg des PaO2 von 25–30 auf 90–95 mmHg kurz nach der Geburt kommt es zu einem Absinken des Serumerythropoetinspiegels, was wiederum zur Folge hat, dass die Bildung der Erythrozyten zwischen der Geburt und dem Alter von 6–8 Wochen sistiert. Es kommt zur physiologischen Anämie bei Reifgeborenen bzw. zur Frühgeborenenanämie. Dieser physiologische Rückgang der zirkulierenden roten Blutkörperchen regt die Produktion von roten Blutkörperchen im Knochenmark an und erfordert in der Regel keine Behandlung.
Neonatale hämatopoetische Funktion Literatur
1. Aher S, Malwatkar K, Kadam S: Neonatal anemia. Semin Fetal Neonatal Med 13(4):239–247, 2008. doi: 10.1016/j.siny.2008.02.009
Neonatale immunologische Funktion
(Siehe auch Zelluläre Komponenten des Immunsystems und Molekulare Komponenten des Immunsystems.)
Die Immunfunktion des Fetus entwickelt sich im Laufe der Schwangerschaft, und daher ist die Immunschwäche bei Frühgeborenen, die in einem früheren Schwangerschaftsalter geboren werden, am schwersten. Die meisten Immunmechanismen sind selbst bei voll ausgetragenen Neugeborenen noch nicht voll funktionsfähig. Daher ist die Funktion des Immunsystems bei allen Neugeborenen und jungen Säuglingen im Vergleich zu Erwachsenen schwächer ausgebildet, so dass ein erhöhtes Risiko für schwere Infektionen besteht. Dieses Risiko wird durch mütterliche Erkrankungen, neonatalen Stress und Medikamente (z. B. Immunsuppressiva, Antiepileptika) erhöht. Die reduzierte Immunantwort des Neugeborenen erklärt auch das Fehlen von Fieber oder lokaler klinischer Krankheitszeichen (z. B. Meningismus) der Infektion.
Die im Dottersackstadium bereits erstmals vorhandenen phagozytierenden Zellen sind während der Fetalzeit maßgeblich für die antiinflammatorische Antwort bei der Abwehr von Bakterien- und Pilzinfektionen verantwortlich. Granulozyten können in der 2. Schwangerschaftsmonat und Monozyten im 4. Schwangersmonat identifiziert werden. Ihre Funktionsfähigkeit verbessert sich mit zunehmendem Gestationsalter, ist zum Geburtstermin aber immer noch eingeschränkt.
Die Ultrastruktur der neutrophilen Granulozyten ist bei Geburt zwar normal, allerdings ist die Chemotaxis der Neutrophilen und Monozyten bei den meisten Neugeborenen aufgrund intrinsischer Fehlfunktionen der Zellbeweglichkeit und der Adhäsionsfähigkeit vermindert. Bei unreifen Kindern ist diese Beeinträchtigung der Funktion noch ausgeprägter.
Der Thymus funktioniert ungefähr ab der 14. Schwangerschaftswoche und durch Blut bildende Zellen produzierte Lymphozyten sammeln sich zur Entwicklung im Thymus an. Ebenfalls mit der 14. Woche sind in der fetalen Leber und Milz T-Zellen vorhanden, was darauf hindeutet, dass in diesem Alter erstmals reife T-Zellen in den sekundären peripheren lymphatischen Organen vorhanden sind. Der Thymus zeigt seine größte Aktivität während der fetalen Entwicklung. Intrauterin weist er ein starkes Wachstum auf und ist beim gesunden Neugeborenen auf dem Röntgen-Bild des Thorax leicht zu erkennen. Die maximale Größe wird mit dem 10. Lebensjahr erreicht, von da an bildet sich der Thymus über viele Jahre langsam zurück.
Die Anzahl der T-Lymphozyten im fetalen Kreislauf nimmt während des 2. Trimesters kontinuierlich zu und erreicht in der 30.–32. Schwangerschaftswoche Normalwerte. Unmittelbar nach der Geburt haben Säuglinge im Vergleich zu Erwachsenen eine relative Lymphozytose. Die Funktionalität der neonatalen T-Lymphozyten ist geringer als die der adulten T-Lymphozyten. So kann es zu einer unzureichenden Immunantwort auf Antigene und zu einer unzureichenden Produktion von Zytokinen kommen.
B-Lymphozyten können in der 12. Schwangerschaftswoche im fetalen Knochenmark, Blut, Leber und Milz nachgewiesen werden. Bis zur 20. Woche können Spuren von IgM und IgG und bis zur 30. Woche Spuren von IgA nachgewiesen werden. Da sich der Fetus normalerweise in einer antigenfreien Umgebung befindet, werden in utero nur kleine Mengen Immunglobuline (vorwiegend IgM) gebildet. Erhöhte IgM-Spiegel im Nabelschnurblut weisen auf eine intrauterine Auseinandersetzung mit Antigenen hin, meistens durch eine konnatale Infektion. Fast das gesamte IgG wird diaplazentar von der Mutter erworben. Der plazentare Transfer des IgG erhöht sich ab der 22. Gestationswoche, bei der Geburt erreicht oder übersteigt die IgG-Konzentration mütterliche Werte. Bei Frühgeborenen liegt die IgG-Konzentration entsprechend dem Ausmaß der Unreife niedriger.
Der passive Transfer der mütterlichen Immunität durch transplazentares IgG und sekretorisches IgA sowie antimikrobielle Faktoren in der Muttermilch (z. B. IgG, sekretorisches IgA, Leukozyten, Komplementfaktoren, Lysozyme und Lactoferrin) kompensiert das unreife Immunsystem des Neugeborenen und verleiht eine Immunität gegen viele Bakterien und Viren. Protektive Immunfaktoren aus der Muttermilch kleiden den Darm und die oberen Luftwege in Verbindung mit dem schleimhautassoziierten lymphatischen Gewebe aus und verringern damit die Wahrscheinlichkeit des Befalls der Schleimhäute mit respiratorischen und intestinalen Pathogenen.
Mit der Zeit nimmt die passive Immunität ab und erreicht im Alter von drei bis sechs Monaten ihr Minimum. Insbesondere Frühgeborene können in den ersten 6 Lebensmonaten eine ausgeprägte Hypogammaglobulinämie aufweisen. Im ersten Lebensjahr liegt der IgG-Spiegel bei etwa 60% des Erwachsenenwertes. Die Konzentration von IgA, IgM, IgD und IgE, die die Plazenta nicht passieren und daher bei Geburt nur in Spuren vorhanden sind, nehmen im Laufe der Kindheit langsam zu. Die IgA-, IgM- und IgA-Spiegel entsprechen im Alter von etwa 10Jahren denen eines Erwachsenen.
Obwohl die Antikörperreaktion auf die ersten Impfdosen bei Frühchen geringer ausfällt als bei termingerechten Säuglingen, sind Frühchen immer noch in der Lage, eine Schutzreaktion auf die meisten Impfstoffe auszulösen, und sollten nach demselben Zeitplan geimpft werden wie termingerechte Säuglinge. Säuglinge, die zum Zeitpunkt der ersten Dosis des Hepatitis-B-Impfstoffs < 2 kg wiegen, sollten jedoch 3 zusätzliche Dosen erhalten, wenn die erste Dosis im Alter von < 1 Monat verabreicht wird weil sie eine verminderte Antikörperreaktion aufweisen (1).
Literatur zur neonatalen immunologischen Funktion
1. Gagneur A, Pinquier D, Quach C: Immunization of preterm infants. Hum Vaccin Immunother 11(11):2556–2563, 2015. doi: 10.1080/21645515.2015.1074358
Neonatale Lungenfunktion
Die fetale Lungenentwicklung vorläuft durch Phasen der Organogenese und Differenzierung. Ziemlich gut entwickelte Alveolen und Typ-II-Tensid herstellende Pneumozyten liegen um die 25. Woche vor und reifen auch weiterhin während der gesamten Gestation. Die kontinuierlich von der Lunge produzierte Flüssigkeit setzt sich aus einem Transsudat aus den Lungenkapillaren und dem Surfactant-Faktor, der von den Typ-II-Pneumozyten sezerniert wird, zusammen. Damit bei der Geburt ein normaler Gasaustausch stattfinden kann, müssen die pulmonale Alveolarflüssigkeit und die interstitielle Flüssigkeit umgehend geklärt werden. Dieser Ausscheidungsprozess erfolgt in erster Linie durch die Aufnahme von Flüssigkeit in die Zellen der Lunge über die Aktivierung von Natriumkanälen im Epithel. Die Kompression des fetalen Thorax während der Geburt trägt nur wenig zur Ausscheidung von Lungenflüssigkeit bei (1). Die transiente Tachypnoe des Neugeborenen wird wahrscheinlich durch eine Verzögerung dieser Mechanismen verursacht.
Bei der Geburt sorgen die elastischen Rückstellkräfte der Rippen und die ersten kräftigen Atembewegungen dafür, dass Luft in den Bronchialbaum eingesogen wird und eine Luft-Wasser-Grenzschicht in den Alveolen entsteht. Beim ersten Atemzug wird Surfactant in diese Grenzschicht freigesetzt. Surfactant, eine Mischung aus Phospholipiden (Phosphatidylcholin, Phosphatidylglycerol, Phosphatidylinositol), neutralen Lipiden und vier oberflächenaktiven Proteinen, die alle in lamellären Einschlusskörperchen der Typ-II-Pneumozyten gespeichert sind, reduziert die Oberflächenspannung, die andernfalls Atelektasen und eine vermehrte Atemarbeit zur Folge hätte. Surfactant wirkt effektiver in kleinen Alveolen als in großen Alveolen, also gegen die normale Tendenz, dass die kleinen Alveolen zu großen Alveolen kollabieren (gemäßg dem Laplace-Gesetz, das besagt, dass in einem elastischen Hohlraum der Druck sinkt während das Volumen zunimmt).
Bei einigen Neugeborenen wird Surfactant eventuell nicht in ausreichenden Mengen produziert, um diffuse Atelektasen wirksam zu verhindern, so dass sich dann ein Atemnotsyndrom entwickelt. Die Produktion und Funktion von Surfactant kann vermindert werden durch Frühgeburt, mütterlicher Diabetes, neonatale Mekoniumaspiration, und neonatale Sepsis. Die neonatale Tensid-Produktion bei Frühgeborenen kann erhöht werden, indem 24 bis 48 h vor der Entbindung Kortikosteroide an die Mutter gegeben werden. Intratracheales Surfactant kann dem Neugeborenen auch nach der Geburt verabreicht werden.
Literatur zur neonatalen Lungenfunktion
1. Ramachandrappa A, Jain L: Elective cesarean section: Its impact on neonatal respiratory outcome. Clin Perinatol 35(2):373–393, vii, 2008. doi: 10.1016/j.clp.2008.03.006
Neonatale Nierenfunktion
Bei der Geburt ist die Nierenfunktion v. a. bei Frühgeborenen in der Regel vermindert.
Die glomeruläre Filtrationsrate steigt mit zunehmendem Gestationsalter kontinuierlich an, v. a. während des 3. Trimesters. Die glomeruläre Filtrationsrate steigt in den ersten Monaten des Lebensschnell, jedoch erreichen glomeruläre Filtrationsrate, Harnstoff-Clearance und maximale tubuläre Clearance bis zum Alter von 1–2 Jahren die Werte von Erwachsenen nicht.