




Das Komplementsystem stellt eine Kaskade von Enzymen dar, die bei der Abwehr von Infektionen zusammenwirken. Viele Komplementproteine sind im Serum als inaktive Enzymvorstufen zufinden (Zymogene); andere befinden sich auf Zelloberflächen. (Siehe auch Übersicht über das Immunsystem.)
Das Komplementsystem überbrückt die angeborene und erworbene Immunität durch
Eine Verstärkung der Antikörper (Ak)-Antworten und des immunologischen Gedächtnisses
Lyse-fremde Zellen
Clearin-Immunkomplexe und apoptotische Zellen
Die Komplementkomponenten erfüllen zahlreiche biologische Funktionen (z. B. Stimulation der Chemotaxis, Auslösen der Mastzelldegranulation unabhängig von Immunoglobulin E).
Komplementaktivierung
Es gibt 3 Wege der Komplementaktivierung (siehe Abbildung Wege der Komplementaktivierung):
Klassisch
Lektin
Alternativ
Wege der Komplementaktivierung
Der klassische, der Lektin- und der alternative Weg münden zuletzt in einem gemeinsamen Weg, wenn die C3-Konvertase (C3 con) C3 in C3a und C3b spaltet. Ab = Antikörper; Ag = Antigen; C1-INH = C1-Inhibitor, MAC = Membran-Angriffs-Komplex; MASP = MBL-assoziierten Serinprotease; MBL = Mannose-bindende Lektin. Querstrich zeigt die Aktivierung. |

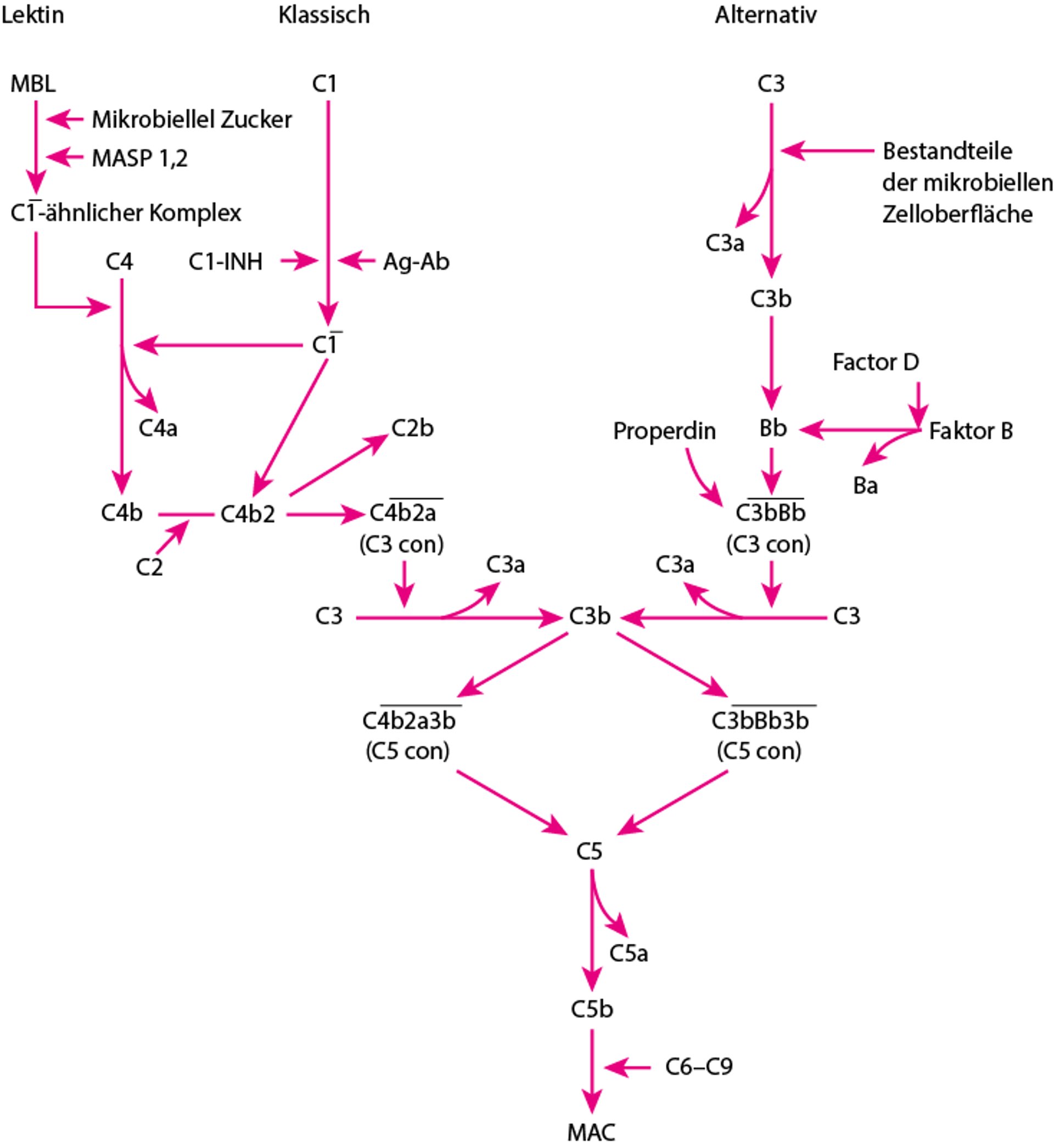
Die Komponenten des klassischen Weges sind – ausgehend von der Reihenfolge, in der sie identifiziert wurden – mit einem C und einer Zahl gekennzeichnet (z. B. C1, C3). Komponenten des alternativen Weges werden oft mit einem Buchstaben (z. B. Faktor B, Faktor D) oder mit einem Namen (z. B. Properdin) versehen.
Der klassischer Weg der Aktivierung ist entweder
Antikörper-abhängig, tritt auf, wenn C1 mit Antigen-IgM oder aggregierten Antigen-IgG-Komplexen interagiert
Antikörper-unabhängig und ereignet sich, wenn Polyanionen (z. B. Heparin, Protamine, DNA und RNA von apoptotischen Zellen), gramnegative Bakterien oder gebundenes C-reaktives Protein (CRP) direkt mit C1 regieren
Dieser Weg wird durch den C1-Inhibitor (C1-INH) reguliert. Das hereditäre Angioödem entsteht aufgrund eines genetischen Mangels an C1-INH.
Die Aktivierung durch den Lektinweg ist Ak-unabhängig; sie entsteht aufgrund der Anbindung von Mannose-bindendem Lektin (MBL), einem Serumprotein, an Mannose, Frukose, oder N-Azetylglukosamin-Gruppen auf bakteriellen Zellwänden, Hefezellwänden oder Viren. Ansonsten gleicht dieser Weg strukturell und funktionell dem klassischen Weg.
Die Aktivierung des alternativen Weges wird eingeleitet, wenn Komponenten mikrobieller Zelloberflächen (z. B. Hefezellwände, Lipopolysaccharide [Endotoxin] der Bakterienzellwände) oder Immunglobulin (z. B. nephritischer Faktor, aggregiertes IgA) kleine Mengen C3 spalten. Dieser Weg wird durch Properdin, Faktor H und DAF (decay accelerating factor, CD55) reguliert.
Die drei Aktivierungswege münden zuletzt in einen gemeinsamen Weg, wenn die C3-Konvertase C3 in C3a und C3b spaltet (siehe Abbildung Wege der Komplementaktivierung). Die C3-Spaltung kann zur Bildung des Membranangriffskomplexes (MAK), der zytotoxischen Komponente des Komplementsystems, führen. MAK veranlasst die Lyse von Fremdzellen.
Faktor I mit Kofaktoren, einschließlich Membran-Kofaktor-Protein (CD46), deaktiviert C3b und C4b.
Komplementärmängel und Defekte
Mängel oder Defekte bei spezifischen Komplementkomponenten wurden mit spezifischen Störungen in Verbindung gebracht; Die folgenden sind Beispiele:
Mangel an C1, C2, C3, MBL, MBL-assoziierten Serinproteas (MASP-2), Faktor H, Faktor I oder Komplementrezeptor 2 (CR2): Anfälligkeit für wiederkehrende bakterielle Infektionen
Mangel an C5, C9, Faktor B, Faktor D, oder Properdin: Anfälligkeit für Neisseria-Infektionen
Mängel bei C1, C4 und C5: systemischem Lupus erythematodes
Defects bei CR2: Allgemeine variable Immundefizienz
Mängel von CR3Leukozytenadhäsionsdefekt Typ 1
Mutationen in den Genen für Faktor B, Faktor H, Faktor I, Membran-Cofaktor-Protein (CD46) oder C3: Entwicklung der atypischen Variante von hämolytisch-urämisches Syndrom
Biologische Aktivitäten der Ergänzung
Komplementkomponenten erfüllen auch andere Immunfunktionen. Diese werden durch Komplementrezeptoren (CR) vermittelt, die sich auf verschiedenen Zellen befinden. Mehrere CRs verwenden Moleküle, denen eine CD-Nummer zugewiesen wurde.
CR1 (CD35) unterstützt die Phagozytose und hilft bei der Auflösung der Immunkomplexe.
CR2 (CD21) reguliert die Antikörper-Produktion durch B-Zellen und ist der Rezeptor des Epstein-Barr-Virus.
CR3 (CD11b/CD18), CR4 (CD11c/CD18) und C1q-Rezeptoren spielen eine Rolle bei der Phagozytose.
C3a, C5a und C4a (schwach) besitzen Anaphylatoxinaktivität: Sie verursachen die Degranulation der Mastzelllen, was zu erhöhter Gefäßpermeabilität und Kontraktion der glatten Muskulatur führt.
C3b fungiert als Opsonin durch Ummantelung der Mikroorganismen und der daraus resultierenden Unterstützung der Phagozytose.
C3d verbessert die Antikörperproduktion durch B-Zellen.
C5 ist ein chemischer Lockstoff, der die Aktivität von Neutrophilen und Monozyten reguliert und die Adhärenz der Zellen steigern, eine Degranulation und Freisetzung intrazellulärer Enzyme aus Granulozyten initiieren, zur Produktion toxischer Sauerstoffmetabolite führen sowie andere metabolische Ereignisse in den Zellen bewirken kann.